

Abstract
Dysregulation of cancer cell bioenergetics is one of the hallmarks of cancer. The Warburg effect is one such documented change. However, glucose metabolism is not universally increased in cancer cells. Uptake of radiolabelled glucose in chronic lymphocytic leukaemia (CLL) fails as a marker of proliferation and the underlying reason is not well elucidated [Conte et al, 2014]. Using proteomic and comprehensive lipid analyses, the preferred metabolic pathways of CLL cells have been identified.
We have previously shown by proteomic analysis that circulating peripheral blood CLL cells demonstrate a significant increase in the expression of proteins pivotal in endogenous lipid synthesis pathways compared to B-cells from healthy individuals. These include fatty acid synthase (+3.84 fold), farnesyl diphosphate synthase (+2.17 fold), ATP-citrate lyase (+4.63 fold), citrate synthase (+2.45 fold) and acetyl-CoA acetyltransferase 1 (+5.84 fold). Conversely, analysis of the proliferation centres in CLL lymph nodes reveals increased expression of proteins involved in beta-oxidation, such as hydroxyacyl-CoA dehydrogenase (+2.02 fold). Additionally CLL cells shows a large increase in the levels of phospholipids found associated with lipid droplets compared to healthy B-cells, including phosphatidylinositol and phosphatidylethanolamine.
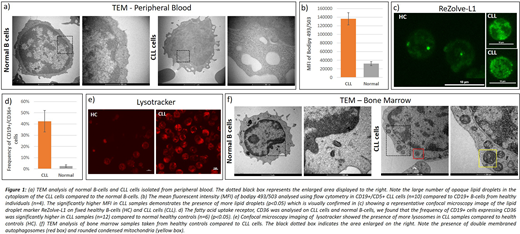
Our current studies have expanded these observations in an attempt to determine if CLL cells have a preponderance for endogenous synthesis of lipids or exogenous uptake. We performed a comprehensive analysis of CLL cells from the bone marrow and peripheral blood using a variety of techniques. Transmission electron microscopy (TEM) images demonstrate striking morphological differences between normal B-cells and CLL cells with the inclusion of lipid droplets in the cytoplasm of CLL cells (Figure 1A) which was most evident in peripheral CLL samples. These were confirmed to be lipid droplets by the specific marker Bodipy 493/503 on flow cytometry (Fig. 1B) and confocal microscopy imaging with ReZolveL1 which targets neutral lipids (Fig. 1C). These showed a substantially higher level of lipid droplet staining in CLL cells compared to normal B-cells. Additionally there was higher expression of CD36 in CLL cells which is the receptor for exogenous lipid uptake into cells (Fig 1D).
TEM analysis also revealed a high number of lysosomes in CLL cells, which was confirmed with lysotracker imaging on confocal microscopy (Fig. 1E). Lysosomes are now emerging as key players in lipid transport and biogenesis [Thelen and Zoncu, 2017]. Lipid droplets may be broken down by a process called autophagy mediated lipid degradation, or lipophagy, which relies on autophagosomes delivering lipid droplets to lysosomes. Autophagosomes were observed in several further bone marrow samples (Fig. 1F). The bone marrow samples also showed evidence of 'condensed mitochondria', indicative of high rates of beta-oxidation [Rossignol et al, 2004].
qPCR was then carried out on 90 genes involved in lipid metabolism. These results revealed significant differences between healthy B-cells and CLL cells. All CLL samples analysed demonstrated high expression of PLIN1, which protects lipid droplets from degradation, and low expression of APOC1, which prevents fatty acid uptake. From these observations we propose that peripheral CLL cells scavenge lipids from the periphery, mainly by uptake via the CD36 receptor, and increased endogenous lipid synthesis pathways.
These results underpin a model for metabolism and enhanced survival for CLL cells in the hypoxic and potentially nutrient poor bone marrow/lymph node microenvironments. Peripheral CLL cells scavenge lipids; these excess lipids, which are toxic to the cell, are stored in lipid droplets which are protected from degradation by a high expression of PLIN proteins. Once these cells circulate back to the bone marrow and lymph nodes to proliferate, the lipid droplets are degraded, likely by lipophagy and neutral lipolysis which frees fatty acids to be used in beta oxidation in the mitochondria to sustain cell proliferation in the bone marrow and lymph node compartments. Our results begin to unravel CLL bioenergetics and dysregulation of cellular metabolism that occurs in this disease. We are now investigating whether the manipulation of these pathways, particularly lipophagy, may represent a novel therapeutic approach in CLL.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal